
![]() This post was originally written in 2006 and re-posted a few times, including in 2010.
This post was originally written in 2006 and re-posted a few times, including in 2010.
As you may know, I have been teaching BIO101 (and also the BIO102 Lab) to non-traditional students in an adult education program for about twelve years now. Every now and then I muse about it publicly on the blog (see this, this, this, this, this, this and this for a few short posts about various aspects of it – from the use of videos, to the use of a classroom blog, to the importance of Open Access so students can read primary literature). The quality of students in this program has steadily risen over the years, but I am still highly constrained with time: I have eight 4-hour meetings with the students over eight weeks. In this period I have to teach them all of biology they need for their non-science majors, plus leave enough time for each student to give a presentation (on the science of their favourite plant and animal) and for two exams. Thus I have to strip the lectures to the bare bones, and hope that those bare bones are what non-science majors really need to know: concepts rather than factoids, relationship with the rest of their lives rather than relationship with the other sciences. Thus I follow my lectures with videos and classroom discussions, and their homework consists of finding cool biology videos or articles and posting the links on the classroom blog for all to see. A couple of times I used malaria as a thread that connected all the topics – from cell biology to ecology to physiology to evolution. I think that worked well but it is hard to do. They also write a final paper on some aspect of physiology.
Another new development is that the administration has realized that most of the faculty have been with the school for many years. We are experienced, and apparently we know what we are doing. Thus they recently gave us much more freedom to design our own syllabus instead of following a pre-defined one, as long as the ultimate goals of the class remain the same. I am not exactly sure when am I teaching the BIO101 lectures again (late Fall, Spring?) but I want to start rethinking my class early. I am also worried that, since I am not actively doing research in the lab and thus not following the literature as closely, that some of the things I teach are now out-dated. Not that anyone can possibly keep up with all the advances in all the areas of Biology which is so huge, but at least big updates that affect teaching of introductory courses are stuff I need to know.
I need to catch up and upgrade my lecture notes. And what better way than crowdsource! So, over the new few weeks, I will re-post my old lecture notes (note that they are just intros – discussions and videos etc. follow them in the classroom) and will ask you to fact-check me. If I got something wrong or something is out of date, let me know (but don’t push just your own preferred hypothesis if a question is not yet settled – give me the entire controversy explanation instead). If something is glaringly missing, let me know. If something can be said in a nicer language – edit my sentences. If you are aware of cool images, articles, blog-posts, videos, podcasts, visualizations, animations, games, etc. that can be used to explain these basic concepts, let me know. And at the end, once we do this with all the lectures, let’s discuss the overall syllabus – is there a better way to organize all this material for such a fast-paced class.
Adaptation vs. Diversity
Biology is concerned with answering two Big Questions: how to explain the adaptation of organisms to their environments and how to explain the diversity of life on Earth.
Much of the course content so far engaged the question of the origin and evolution of adaptation, and much of the remainder of the course will also look at particular adaptations of humans and other vertebrates. This is the only lecture specifically targeting the question of diversity.
The way this material is usually taught is to go over long lists of organisms and tabulate their characters, how the members of one group are similar to each other and different from members of other groups. We, in our course, will try a different approach, i.e., not just describing, but also explaining diversity – how it comes about.
If you think about it, knowing what we learned so far about the way evolution works, the origin of adaptation and the origin of diversity are deeply intertwined: as local populations evolve adaptations to their current local environments, they become more and more different from each other until the species splits into two or more new species. Thus, evolution of adaptations to local conditions leads to proliferation of new species, thus to the increase in overall diversity of life on the planet.
Origin of Life
One can postulate four ways the life on Earth came about: a) it was created – poof! – out of nothing by an intelligent being, e.g., God; b) it was created – poof! – out of nothing by an intelligent being, e.g., space aliens, either on Earth or elsewhere, then brought to Earth; c) it spontaneously arose elsewhere in the Universe and was brought to Earth by comets and meteors; and d) it spontaneously arose out of chemical reactions in the ancient seas in the presence of the ancient atmosphere.
Science is incapable of addressing the first notion – being untestable and unfalsifiable (impossible to prove that it is wrong), it is properly outside of the realm of science and within the domain of religion.
The first three notions also just move the goalposts one step further – how did life (including God and/or Aliens) arise elsewhere in the Universe? Thus, scientists focus only on the one remaining testable hypothesis – the one about spontaneous and gradual generation of life out of non-life, a process called abiogenesis. The scientific study of abiogenesis cannot say and does not attempt to say, anything about existence of God or Aliens. It only attempts to figure out how life could have arisen on its own, sometime between 3 and 4 billion years ago.
All of life on Earth descends from a single common ancestor. It is quite possible that life initially arose multiple times, but as soon as one life form became established and competitive enough, all the other instances of spontaneous generation of life were out-competed and did not leave progeny.
It is difficult to study the origin of life as molecules do not leave fossils. They do leave chemical traces, though, so we know a lot about the chemistry of the ancient oceans, soil and atmosphere. Thus, we know under what conditions and what available materials (and energy) life first arose. By replicating such conditions in the laboratory, we can study the details of how life might have evolved out of non-life.
The study of the origin of life is a lively and exciting area of biology, perhaps because so little has yet been settled with great certainty. There are a number of competing hypotheses promoted by various research groups. Those hypotheses can be classified into groups: RNA First, Protein First, RNA-Protein First and Bubbles First.
RNA is a molecule that can be replicated and thus can serve as the original hereditary material (DNA is too large and complex even for some of today’s viruses, let alone for the first simple organisms). RNA is also capable of catalytic activity – promoting and speeding up reactions between other molecules, as well as replicating itself. Thus, RNA is the best candidate for the first molecule of life. Still, it is not capable of everything that life needs, so a few simple polypeptides (and those are really easy to synthesize in a flask mimicking the original Earth conditions) were probably involved from the very beginning. For those reactions to occur, they had to be separated from the remaining ocean – thus some kind of “cell membrane”, like a soap bubble, was also necessary for the origin and early evolution of life.
Those early “cells” competed against each other. Those that, through chemical evolution, managed to become good enough at remaining stable for a decent amount of time, capable of acquiring the energy from the environment, and capable of dividing into two “daughter cells” out-competed the others – chemical evolution turned into biological evolution. As they changed through trial and error, some cells gradually got better at “living” and out-competed all others. One best competitor of the early living world is the common ancestor of all of the subsequent life on Earth, including us.
Directionality of Evolution
There are two common misconceptions about evolution. First is the idea that evolution tends towards perfection. But, always remember that evolution favors individuals who are slightly better optimized to current local conditions than other individuals of the same species, i.e., what wins is the relative fitness, not absolute fitness (i.e., perfection). In other words, you have to be capable of surviving and reproducing in your current environment and be just a tad little bit better at it than your conspecifics – there is no need to be perfectly adapted.
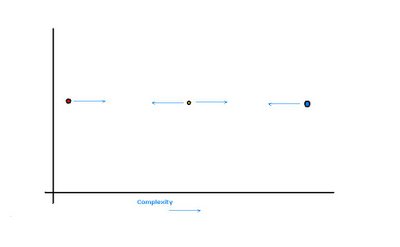
The second common misconception about evolution is that it has a tendency to generate greater complexity. Originally, right after the initial origin of life on Earth, evolution did produce greater complexity, but only because there was no way to become any more simple than the first organisms already were. There is a “left wall” of complexity in the living world, i.e. there is a minimum complexity that is necessary for something to be deemed alive.
 Thus, initially, the only direction evolution could take was away from the left wall (red dot), i.e., becoming more complex. But once reasonably complex organisms evolved, they were not snuggled against the left wall any more (yellow dot). Adaptation to current local conditions can equally promote simplification as it does complexification of the organism in question. In other words, as populations evolve, the members of the populations are equally likely to become simpler as they are to become more complex.
Thus, initially, the only direction evolution could take was away from the left wall (red dot), i.e., becoming more complex. But once reasonably complex organisms evolved, they were not snuggled against the left wall any more (yellow dot). Adaptation to current local conditions can equally promote simplification as it does complexification of the organism in question. In other words, as populations evolve, the members of the populations are equally likely to become simpler as they are to become more complex.
Actually, as we know from the world of man-made machines, there is such a thing as being too complex (blue dot). Over-complicated machines break down much more easily and are more difficult to maintain and repair. Likewise, organisms of great complexity are often not as fit as their simpler relatives – their genomes are so large that the error rate is greater and cell division is more difficult. Cells can “go wild” and turn into cancer. Also, with so many interacting parts, it is more difficult for complicated organisms to evolve new adaptations as the development of the whole complex system has to change and adapt to such changes.
Thus, simplification is as often seen in evolution as is acquisition of greater complexity. Just think of parasites – they are all simplified versions of their free-living relatives – no need for eyes, other sensory organs or means of locomotion if one spends one’s life attached to the lining of the host’s intestine, sucking in nutrients and growing billions of eggs.
Measuring Diversity – Taxonomy and Systematics
People have always tried to classify living beings around them, by grouping them according to some man-made criteria, usually by the way they look, where they live, and how useful they may be to us. Only for the past 150 years we have understood that all organisms on our planet are genealogically related to each other, so we started classifying them according to the degree of relatedness – drawing family trees of Life.
Initially, classification was done according to anatomy and embryology of organisms. Such methods brought about the division of Life on Earth into six great Kingdoms: Bacteria, Archaea, Protista, Plants, Fungi and Animals. The first two are Prokaryotes (cell has no nucleus), the latter four are Eukaryotes (cells have a nucleus).
The Kingdoms were, like Russian dolls, further subdivided into nested hierarchies: each Kingdom was composed of a number of Phyla (Phylum = type). Each Phylum consists of Classes, those are made of Orders that are further subdivided into Families. Each family consists of Genera and each Genus is composed of the most closely related Species.
The proper name of each living organism on Earth is its binomial Latin name – capitalized name of the Genus and lower-case name of the species, both italicized, e.g., Homo sapiens, Canis familiaris, Equus caballus, Bos taurus (human, dog, horse and cow, respectively).
Lately, modern molecular genetic techniques have been applied to testing relationships between species, resulting in many changes in classification at lower levels of systematics (e.g,. species, genus, family, etc).
The knowledge gained from this approach also resulted in some big changes in the way we classify living organisms. Instead of six Kingdoms, we now divide life on Earth into three Domains: Bacteria, Archaea and Eukarya.
We are now aware that endosymbiosis (intercellular parasites, originally small bacterial cells entering and living inside larger bacterial cells) gave rise to organelles, like mitochondria and chloroplasts. We are now aware how much lateral (or horizontal) transfer of genetic material is going on between species, i.e., the branching tree of life has many traversing connections between branches as well.
Cladistics is a relatively new method of classifying organisms, using multiple (often many) different genetic, morphological and other traits and building “trees” by calculating (using computer software) the probabilities of each two of the species being related to each other. Thus, “most likely” trees are plotted as cladograms which can further be tested and refined.
Previously in this series:
BIO101 – Biology and the Scientific Method
BIO101 – Cell Structure
BIO101 – Protein Synthesis: Transcription and Translation
BIO101: Cell-Cell Interactions
BIO101 – From One Cell To Two: Cell Division and DNA Replication
BIO101 – From Two Cells To Many: Cell Differentiation and Embryonic Development
BIO101 – From Genes To Traits: How Genotype Affects Phenotype
BIO101 – From Genes To Species: A Primer on Evolution
BIO101 – What Creatures Do: Animal Behavior
BIO101 – Organisms In Time and Space: Ecology










{kind=link}