
![]() This post was originally written in 2006 and re-posted a few times, including in 2010.
This post was originally written in 2006 and re-posted a few times, including in 2010.
As you may know, I have been teaching BIO101 (and also the BIO102 Lab) to non-traditional students in an adult education program for about twelve years now. Every now and then I muse about it publicly on the blog (see this, this, this, this, this, this and this for a few short posts about various aspects of it – from the use of videos, to the use of a classroom blog, to the importance of Open Access so students can read primary literature). The quality of students in this program has steadily risen over the years, but I am still highly constrained with time: I have eight 4-hour meetings with the students over eight weeks. In this period I have to teach them all of biology they need for their non-science majors, plus leave enough time for each student to give a presentation (on the science of their favourite plant and animal) and for two exams. Thus I have to strip the lectures to the bare bones, and hope that those bare bones are what non-science majors really need to know: concepts rather than factoids, relationship with the rest of their lives rather than relationship with the other sciences. Thus I follow my lectures with videos and classroom discussions, and their homework consists of finding cool biology videos or articles and posting the links on the classroom blog for all to see. A couple of times I used malaria as a thread that connected all the topics – from cell biology to ecology to physiology to evolution. I think that worked well but it is hard to do. They also write a final paper on some aspect of physiology.
Another new development is that the administration has realized that most of the faculty have been with the school for many years. We are experienced, and apparently we know what we are doing. Thus they recently gave us much more freedom to design our own syllabus instead of following a pre-defined one, as long as the ultimate goals of the class remain the same. I am not exactly sure when am I teaching the BIO101 lectures again (late Fall, Spring?) but I want to start rethinking my class early. I am also worried that, since I am not actively doing research in the lab and thus not following the literature as closely, that some of the things I teach are now out-dated. Not that anyone can possibly keep up with all the advances in all the areas of Biology which is so huge, but at least big updates that affect teaching of introductory courses are stuff I need to know.
I need to catch up and upgrade my lecture notes. And what better way than crowdsource! So, over the new few weeks, I will re-post my old lecture notes (note that they are just intros – discussions and videos etc. follow them in the classroom) and will ask you to fact-check me. If I got something wrong or something is out of date, let me know (but don’t push just your own preferred hypothesis if a question is not yet settled – give me the entire controversy explanation instead). If something is glaringly missing, let me know. If something can be said in a nicer language – edit my sentences. If you are aware of cool images, articles, blog-posts, videos, podcasts, visualizations, animations, games, etc. that can be used to explain these basic concepts, let me know. And at the end, once we do this with all the lectures, let’s discuss the overall syllabus – is there a better way to organize all this material for such a fast-paced class.
Today, we continue with the cell biology portion of the course – covering the way cells communicate with each other, something that will come up over and over again for the rest of the course. See the previous lectures:
Biology and the Scientific Method
BIO101 – Cell Structure
BIO101 – Protein Synthesis: Transcription and Translation
————————————————-
BIO101 – Bora Zivkovic – Lecture 1 – Part 4
Cell-cell interactions
Cells do not exist in complete isolation. For a coordinated function of cells in a tissue, tissues in an organ, organs in a system and systems in the body, cells need to be able to communicate with each other. Each cell should be capable of sending chemical signals to other cells and of receiving chemical signals from oter cells, as well as signals (chemical or other) from its immediate environment.
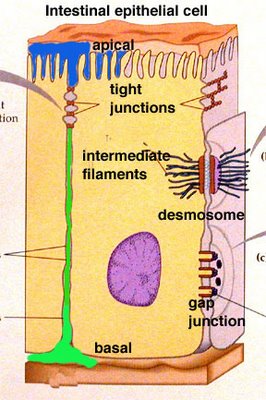
Cell membrane is a double layer of molecules of fat. Some small chemical messengers are capable of passing through the membrane. Most ions and most molecules cannot pass through the membrane, thus the information between the inside and the outside of the cell is mediated by proteins embedded in the membrane.
Membrane proteins serve various functions. For instance, such proteins form tight junctions that serve to glue neighboring cells together and prevent passage of substances between the two cells. Other surface proteins are involved in cell-cell recognition, which is important for the immune response. Other membrane proteins serve functions in communication between the inside of the cell and the cell’s immediate environment.
How does a cell send a signal?
A cell can communicate signals to other cells in various ways. Autocrine signaling is a way for a cell to alter its own extracellular environment, which in turn affects the way the cell functions. The cell secretes chemicals outside of its membrane and the presence of those chemicals on the outside modifies the behavior of that same cell. This process is important for growth.
Paracrine signaling is a way for a cell to affect the behavior of neighboring cells by secreting chemicals into the common intercellular space. This is an important process during embryonic development.
Endocrine signaling utilizes hormones. A cell secretes chemicals into the bloodstream. Those chemicals affect the behavior of distant target cells. We will go into more details of autocrine, paracrine and endocrine signaling later on, when we tackle the human endocrine system.
Direct signaling is a transfer of ions or small molecules from one cell to its neighbor through pores in the membrane. Those pores are built out of membrane proteins and are called gap junctions. This is the fastest mode of cell-cell communication and is found in places where extremely fast and well-coordinated activity of cells in needed. An example of this process can be found in the heart. The muscle cells in the heart communicate with each other via gap junctions which allows all heart cells to contract almost simultaneously.

Finally, synaptic signaling is found in the nervous system. It is a highly specific and localized type of paracrine signalling between two nerve cells or between a nerve cell and a muscle cell. We will go into details of synaptic signaling when we cover the human nervous system.
How does a cell receive a signal?
Some small molecules are capable of entering the cell through the plasma membrane. Nitrous oxide is one example. Upon entering the cell, it activates an enzyme.
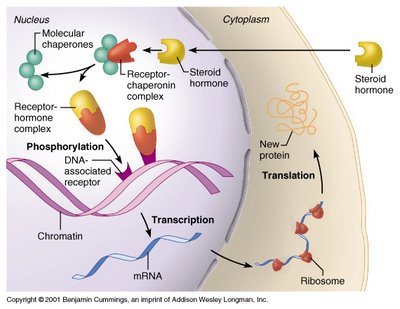
Some small hormones also enter the cell directly, by passing through the membrane. Examples are steroid hormones, thyroid hormones and melatonin. Once inside the cell, they bind cytoplasmic or nuclear receptors. The hormone-receptors complex enters the nucleus and binds to a particular sequence on the DNA. Binding dislodges a protein that inhibits the expression of the gene at that segment, so the gene begins to be transcribed and translated. Thus, a new protein appears in the cell and assumes its normal function within it (or gets secreted). The action of nuclear receptors is slow, as it takes some hours for the whole process to occur. The effect is long-lasting (or even permanent) and changes the properties of the cell. This type of process is important in development, differentiation and maturation of cells, e.g., gametes (eggs and sperm cells).

There are three types of cell surface receptors: membrane enzymes, ion channels, and transmembrane receptors.

When a signaling chemical binds to the membrane enzyme protein on the outside of the cell, this triggers a change in the 3D conformation of that protein, which, in turn, triggers a chemical reaction on the inside of the cell.
When a signaling molecule binds to an ion channel on the outside of the cell, this triggers the change of the 3D conformation of the protein and the channel opens, allowing the ions to move in or out of the cell following their electrical gradients and thus altering the polarization of the cell membrane. Some ion channels respond to non-chemical stimuli in the same way, including changes in electrical charge or mechanical disturbance of the membrane.
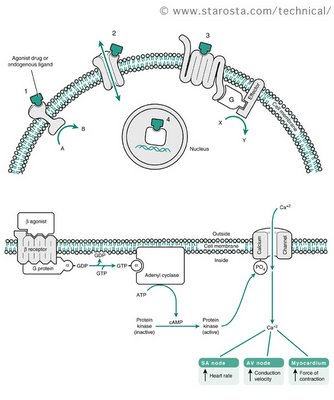
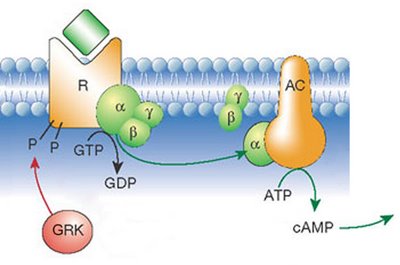
 G protein-linked receptors are seven-pass transmembrane proteins. This means that the polypeptide chain traverses the membrane seven times. When a chemical – a hormone or a pharmaceutical agent – binds to the receptor on the outside of the cell, this triggers a series of chemical reactions, including the movement and binding of the G-protein, transformation of GTP into GDP and activation of second messengers. Second messengers (e.g., cyclic AMP) start a cascade of enzymatic reactions leading to the cellular response. This signaling method is quite fast and, more importantly, it amplifies the signal. Binding of a single hormone molecule quickly results in thousands of molecules of second messengers acting on even more molecules of enzymes and so on. Thus, the response to a small stimulus can be very large. We will go into details of G-protein-mediated signaling when we tackle the endocrine and the sensory systems.
G protein-linked receptors are seven-pass transmembrane proteins. This means that the polypeptide chain traverses the membrane seven times. When a chemical – a hormone or a pharmaceutical agent – binds to the receptor on the outside of the cell, this triggers a series of chemical reactions, including the movement and binding of the G-protein, transformation of GTP into GDP and activation of second messengers. Second messengers (e.g., cyclic AMP) start a cascade of enzymatic reactions leading to the cellular response. This signaling method is quite fast and, more importantly, it amplifies the signal. Binding of a single hormone molecule quickly results in thousands of molecules of second messengers acting on even more molecules of enzymes and so on. Thus, the response to a small stimulus can be very large. We will go into details of G-protein-mediated signaling when we tackle the endocrine and the sensory systems.

Previously in this series:
Biology and the Scientific Method
BIO101 – Cell Structure
BIO101 – Protein Synthesis: Transcription and Translation










{kind=link}
{kind=link}
{kind=link}
{kind=link}