
![]() In this lecture, as well as in the previous one and the next one, I tackle areas of Biology where I am really weak: origin of life, diversity of life, and taxonomy/systematics. These are also areas where there has been a lot of change recently (often not yet incorporated into textbooks), and I am unlikely to be up-to-date, so please help me bring these lectures up to standards…. This post was originally written in 2006 and re-posted a few times, including in 2010.
In this lecture, as well as in the previous one and the next one, I tackle areas of Biology where I am really weak: origin of life, diversity of life, and taxonomy/systematics. These are also areas where there has been a lot of change recently (often not yet incorporated into textbooks), and I am unlikely to be up-to-date, so please help me bring these lectures up to standards…. This post was originally written in 2006 and re-posted a few times, including in 2010.
As you may know, I have been teaching BIO101 (and also the BIO102 Lab) to non-traditional students in an adult education program for about twelve years now. Every now and then I muse about it publicly on the blog (see this, this, this, this, this, this and this for a few short posts about various aspects of it – from the use of videos, to the use of a classroom blog, to the importance of Open Access so students can read primary literature). The quality of students in this program has steadily risen over the years, but I am still highly constrained with time: I have eight 4-hour meetings with the students over eight weeks. In this period I have to teach them all of biology they need for their non-science majors, plus leave enough time for each student to give a presentation (on the science of their favourite plant and animal) and for two exams. Thus I have to strip the lectures to the bare bones, and hope that those bare bones are what non-science majors really need to know: concepts rather than factoids, relationship with the rest of their lives rather than relationship with the other sciences. Thus I follow my lectures with videos and classroom discussions, and their homework consists of finding cool biology videos or articles and posting the links on the classroom blog for all to see. A couple of times I used malaria as a thread that connected all the topics – from cell biology to ecology to physiology to evolution. I think that worked well but it is hard to do. They also write a final paper on some aspect of physiology.
Another new development is that the administration has realized that most of the faculty have been with the school for many years. We are experienced, and apparently we know what we are doing. Thus they recently gave us much more freedom to design our own syllabus instead of following a pre-defined one, as long as the ultimate goals of the class remain the same. I am not exactly sure when am I teaching the BIO101 lectures again (late Fall, Spring?) but I want to start rethinking my class early. I am also worried that, since I am not actively doing research in the lab and thus not following the literature as closely, that some of the things I teach are now out-dated. Not that anyone can possibly keep up with all the advances in all the areas of Biology which is so huge, but at least big updates that affect teaching of introductory courses are stuff I need to know.
I need to catch up and upgrade my lecture notes. And what better way than crowdsource! So, over the new few weeks, I will re-post my old lecture notes (note that they are just intros – discussions and videos etc. follow them in the classroom) and will ask you to fact-check me. If I got something wrong or something is out of date, let me know (but don’t push just your own preferred hypothesis if a question is not yet settled – give me the entire controversy explanation instead). If something is glaringly missing, let me know. If something can be said in a nicer language – edit my sentences. If you are aware of cool images, articles, blog-posts, videos, podcasts, visualizations, animations, games, etc. that can be used to explain these basic concepts, let me know. And at the end, once we do this with all the lectures, let’s discuss the overall syllabus – is there a better way to organize all this material for such a fast-paced class.
These posts are very old, and were initially on a private-set classroom blog, not public. I have no idea where the images come from any more, though many are likely from the textbook I was using at the time. Please let me know if an image is yours, needs to be attributed or removed. Thank you.
=================
It is impossible to cover all organ systems in detail over the course of just two lectures. Thus, we will stick only to the basics. Still, I want to emphasize how much organ systems work together, in concert, to maintain the homeostasis (and rheostasis) of the body. I’d also like to emphasize how fuzzy are the boundaries between organ systems – many organs are, both anatomically and functionally, simultaneously parts of two or more organ systems. So, I will use an example you are familiar with from our study of animal behavior – stress response – to illustrate the unity of the well-coordinated response of all organ systems when faced with a challenge. We will use our old zebra-and-lion example as a roadmap in our exploration of (human, and generally mammalian) physiology:
So, you are a zebra, happily grazing out on the savannah. Suddenly you hear some rustling in the grass. How did you hear it?
The movement of a lion produced oscillations of air. Those oscillations exerted pressure onto the tympanic membrane in your ears. The vibrations of the membrane induced vibrations in three little bones inside the middle ear, which, in turn, induced vibrations of the cochlea in the inner ear.
 Cochlea is a long tube wrapped in a spiral. If the pitch of the sound is high (high frequency of oscillations), only the first portion of the cochlea vibrates. With the lowest frequences, even the tip of the cochlea starts vibrating. Cochlea is filled with fluid. Withing this fluid there is a thin membrane transecting the cochlea along its length. When the cochlea vibrates, this membrane also vibrates and those vibrations move the hair-like protrusions on the surface of sensory cells in the cochlea. Those cells send electrical impulses to the brain, where the sound is processed and becomes a conscious sensation – you have heard the lion move.
Cochlea is a long tube wrapped in a spiral. If the pitch of the sound is high (high frequency of oscillations), only the first portion of the cochlea vibrates. With the lowest frequences, even the tip of the cochlea starts vibrating. Cochlea is filled with fluid. Withing this fluid there is a thin membrane transecting the cochlea along its length. When the cochlea vibrates, this membrane also vibrates and those vibrations move the hair-like protrusions on the surface of sensory cells in the cochlea. Those cells send electrical impulses to the brain, where the sound is processed and becomes a conscious sensation – you have heard the lion move.

The perception of the sound makes you look – yes, there is a lion stalking you, about to leap! How do you see the lion? The waves of light reflected from the surface of the lion travel to your eyes, enter through the pupil, pass through the lens and hit the retina in the back of the eye.

Photoreceptors in the eye (rods and cones) contain a pigment – a colored molecule – that changes its 3D structure when hit by light. In the rods, this pigment is called rhodopsin and is used for black-and-white vision. In the rods, there are similar pigments – opsins – which are most sensitive to particular wavelengths of light (colors) and are used to detect color. The change in 3D structure of the pigment starts a cascade of biochemical reactions resulting in the changes in the electrical potential of the cell – this information is then transferred to the next cell, the next cell, and so on, until it reaches the brain, where the information about the shape, color and movement of the objects (lion and the surrounding grass) is processed and made conscious.

The ear and the eye are examples of the organs of the sensory system. Hearing is one of many mechanical senses – others include touch, pain, balance, stretch receptors in the muscles and tendons, etc. Many animals are capable of hearing sounds that we cannot detect. For instance, bats and some of their insect prey detect the high-pitched ultrasound (a case of a co-evolutionary arms-race). Likewise for dolphins and some of their fish prey. Dogs do, too – that is why we cannot hear the dog whistle. On the other hand, many large animals, e.g., whales, elephants, giraffes, rhinos, crocodiles and perhaps even cows and horses, can detect the deep rumble of the infrasound.
Vision is a sense that detects radiation in the visible specter. Many animals are capable of seeing light outside of our visible specter. For instance, many insects and birds and some small mammals can see ultraviolet light, while some snakes (e.g., pit vipers like rattlesnakes and boids like pythons) and some insects (e.g., Melanophila beetle and some wasps) can perceive infrared light.
Another type of sense is thermoreception – detection of hot and cold. Chemical senses are attuned to particular molecules. Olfaction (smell) and gustation (taste) are the best known chemical senses. Chemical senses also exist inside of our bodes – they are capable of detecting blood pH, blood levels of oxygen, carbon dioxide, calcium, glucose etc. Finally, some animals are capable of detecting other physical properties of the environment., e.g., the electrical and magnetic fields.
All senses work along the same principles: a stimulus from the external or internal environment is detected by a specialized type of cell. Inside the cell a chemical cascade begins – that is transduction. This changes the properties of the cell – usually its cell membrane potential – which is transmitted from the sensory cell to the neighboring nerve cell, to the next cell, next cell and so on, until it ends in the appropriate area of the nervous system, usually the brain. There, the sum of all stimuli from all the cells of the sensory organ are interpreted (integrated and processed over time) and the neccessary action is triggered. This action can be behavioral (movement), or it can be physiological: maintanance of homeostasis.
The sensory information is processed by the Central Nervous System (CNS): the brain and the spinal cord.

All the nerve cells that take information from the periphery to the CNS are sensory nerves. All the nerves that take the decisions made by the CNS to the effectors – muscles or glands – are motor nerves. The sensory and motor pathways together make Peripheral Nervous System.

The motor pathways are further divided into two domains: somatic nervous system is under voluntary control, while autonomic (vegetative) nervous system is involuntary. Autonomic nervous system has two divisions: sympathetic and parasymphatetic. Symphatetic nervous system is active during stress – it acts on many other organ systems, releasing the energy stores, stimulating organs needed for the response and inhibiting organs of no immediate importance.

Thus, a zebra about to be attacked by a lion is exhibiting stress response. Sympathetic nervous system works to release glucose (energy) stores from the liver, stimulates the organs necessary for the fast escape – muscles – and all the other systems that are needed for providing the muscles with energy – the circulatory and respiratory systems. At the same time, digestion, immunity, excretion and reproduction are inhibited. Once the zebra successfully evades the lion, sympathetic system gets inhibited and the parasympathetic system is stimulated – it reverses all the effects. The two systems work antagonistically to each other: they always have opposite effects.
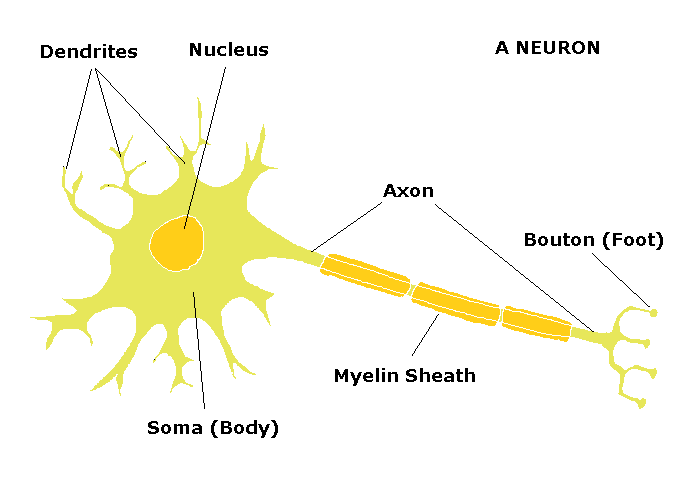
But, how does the nervous system work? Let’s look at the nerve cell – the neuron:

A typical neuron has a cell body (soma) which contains the nucleus and other organelles. It has many thin, short processes – dendrites – that bring information from other neighboring cells into the nerve cell, and one large, long process that takes information away from the cell to another cell – the axon.
There is an electrical potential of the cell membrane – the voltage on the inside and the outside of the cell is different. The inside of the neuron is usually around 70mV more negative (-70mV) compared to the outside. This polarization is accomplished by the specialized proetins in the cell membrane – ion channels and ion transporters. Using energy from ATP, they transport sodium out of the cell and potassium into the cell (also chlorine into the cell). As ions can leak through the membrane to some extent, the cell has to constantly use energy to maintain the resting membrane potential.

An electrical impulse coming from another cell will change the membrane potential of a dendrite. This change is usually not sufficiently large to induce the neuron to respond. However, if many such stimuli occur simultaneously they are additive – the neuron sums up all the stimulatory and inhibitory impulses it gets at any given time. If the sum of impulses is large, the change of membrane potential will still be large when it travels across the soma and onto the very beginning of the axon – axon hillock. If the change of the membrane potential at the axon hillock crosses a threshold (around -40mV or so), this induces sodium channels at the axon hillock to open. Sodium rushes in down its concentration gradient. This results in further depolarization of the membrane, which in turn results in opening even more sodium channels which depolarizes the membrane even more – this is a positive feedback loop – until all of the Na-channels are open and the membrane potential is now positive. Reaching this voltage induces the opening of the potassium channels. Potassium rushes out along its concentration gradient. This results in repolarization of the membrane. The whole process – from initial small depolarization, through the fast Na-driven depolarization, subsequent K-driven repolarization resulting in a small overshoot and the return to the normal resting potential – is called an Action Potential which can be graphed like this:

An action potential generated at the axon hillock results in the changes of membrane potential in the neighboring membrane just down the axon where a new action potential is generated which, in turn, results in a depolarization of the membrane further on down the axon, and so on until the electrical impulse reaches the end of the axon. In vertebrates, special cells called Schwann cells wrap around the axons and serve as isolating tape of sorts. Thus, the action potential instead of spreading gradually down the axon, proceeds in jumps – this makes electrical transmission much faster – something necessary if the axon is three meters long as in the nerves of the hind leg of a giraffe.
What happens at the end of the axon? There, the change of membrane polarity results in the opening of the calcium channels and calcium rushes in (that is why calcium homeostasis is so important). The end of the axon contains many small packets filled with a neurotransmitter. Infusion of calcium stimulates these packets to fuse with the cell membrane and release the neurotransmitter out of the cell. The chemical ends up in a very small space between the axon ending and the membrane of another cell (e.g., a dendrite of another neuron). The membrane of that other cell has membrane receptors that respond to this neurotransmitter. The activation of the receptors results in the local change of membrane potential. Stimulatory neurotransmitters depolarize the membrane (make it more positive), while inhibitory neurotransmitters hyperpolarize the membrane – make it more negative, thus harder to produce an action potential.

The target of a nerve cell can be another neuron, a muscle cell or a gland. Many glands are endocrine glands – they release their chemical products, hormones, into the bloodstream. Hormones act on distant targets via receptors. While transmission of information in the nervous system is very fast – miliseconds, in the endocrine system it takes seconds, minutes, hours, days, months (pregnancy), even years (puberty) to induce the effect in the target. While transmission within the nervous system is local (cell-to-cell) and over very short distances – the gap within a synapse is measured in Angstroms – the transmission within the endocrine system is over long distances and global – it affects every cell that possesses the right kind of receptors.

Many endocrine glands are regulated during the stress response, and many of them participate in the stress response. The thyroid gland releases thyroxine – a hormone that acts via nuclear receptors. Thyroxine has many fuctions in the body and several of those are involved in the energetics of the body – release of energy from the stores and production of heat in the mitochondria. It also produces calcitonin which is one of the regulators of calcium levels in the blood.
Parathyroid gland is, in humans, embedded inside the thyroid gland. Its hormone, parathormone is the key hormone of calcium homeostasis. Calcitonin and parathormone are antagonists: the former lowers and the latter raises blood calcium. Together, they can fine-tune the calcium levels available to neurons, muscles and heart-cells for their normal function.
Pancreas secretes insulin and glucagon. Insulin removes glucose from blood and stores it in muscle and liver cells. Glucagon has the opposite effect – it releases glucose from its stores and makes it available to cells that are in need of energy, e.g., the muscle cells of a running zebra. Together, these two hormones fine-tune the glucose homeostasis of the body.
Adrenal gland has two layers. In the center is the adrenal medulla. It is a part of the nervous system and it releases epineprhine and norepinephrine (also known as adrenaline and noradrenaline). These are the key hormones of the stress response. They have all the same effects as the sympathetic nervous system, which is not surprising as norepinephrine is the neurotransmitter used by the neurons of the sympathetic system (parasympathetic system uses acetylcholine as a transmitter).
The outside layer is the adrenal cortex. It secretes a lot of hormones. The most important are aldosterone (involved in salt and water balance) and cortisol which is another important stress hormone – it mobilizes glucose from its stores and makes it available for the organs that need it. Sex steroid hormones are also produced in the adrenal cortex. Oversecretion of testosterone may lead to development of some male features in women, e.g., growing a beard.
Ovary and testis secrete sex steroid hormones. Testis secretes testosterone, while ovaries secrete estradiol (an estrogen) and progesterone. Progesterone stimulates the growth of mammary glands and prepares the uterus for pregnancy. Estradiol stimulates the development of female secondary sexual characteristics (e.g., general body shape, patterns of fat deposition and hair growth, growth of breasts) and is involved in monthly preparation for pregnancy.
Testosterone is very important in the development of a male embryo. Our default condition is female. Lack of sex steroids during development results in the development of a girl (even if the child is genetically male). Secretion of testosterone at a particular moment during development turns female genitals into male genitals and primes many organs, including the brain, to be responsive to the second big surge of testosterone which happens at the onset of puberty. At that time, primed tissues develop in a male-specific way, developing male secondary sexual characteristics (e.g., deep voice, beard, larger muscle mass, growth of genitalia, male-typical behaviors, etc.).
Many other organs also secrete hormones along with their other functions. The heart, kidney, lung, intestine and skin are all also members of the endocrine system. Thymus is an endocrine gland that is involved in the development of the immune system – once the immune system is mature, thymus shrinks and dissappears.
Many of the endocrine glands are themselves controlled by other hormones secreted by the pituitary gland – the Master Gland of the endocrine system. For instance, the anterior portion of the pituitary gland secretes hormones that stimulate the release of thyroxine from the thyroid gland, cortisol from the adrenal cortex, and sex steroids form the gonads. Other hormones secreted by the anterior pituitary are prolactin (stimulates production of milk, amog else) and growth hormone (which stimulates cells to produce autocrine and paracrine hormones which stimulate cell-division). The posterior portion of the pituitary is actually part of the brain – it secretes two hormones: antidiuretic hormone (control of water balance) and oxytocin (stimulates milk let-down and uterine contractions, among other functions).
All these pituitary hormones are, in turn, controlled (either stimulated or inhibited) by hormones/factors secreted by the hypothalamus which is a part of the brain, which makes the brain the biggest and most important endocrine gland of all.
Pineal organ is a part of the brain (thus central nervous system). In all vertebrates, except mammals and snakes, it is also a sensory organ – it perceieves light (which easily passes through scales/feathers, skin and skull). In seasonally breeding mammals, it is considered to be a part of the reproductive system. In all vertebrates, it is also an endocrine organ – it secretes a hormone melatonin. In all vertebrates, the pineal organ is an important part of the circadian system – a system that is involved in daily timing of all physiological and behavioral functions in the body. In many species of vertebrates, except mammals, the pineal organ is the Master Clock of the circadian system. In mammals, the master clock is located in the hypothalamus of the brain, in a structure known as the suprachiasmatic nucleus (SCN).
Retina is part of the eye (sensory system), it is part of the brain (nervous system), it also secretes melatonin (endocrine system) and contains a circadian clock (circadian system) in all vertebrates. In some species of birds, the master clock is located in the retina of the eye. The day-night differences in light intensity entrain (synchronize) the circadian system with the cycles in the environment. Those differences in light intensity are perceived by the retina, but not by photoreceptor cells (rods and cones). Instead, a small subset of retinal ganglion cells (proper nerve cells) contains a photopigment melanopsin which changes its 3D structure when exposed to light and sends its signals to the SCN in the brain.

Wherever the master clock may be located (SCN, pineal or retina) in any particular species, its main function is to coordinate the timing of peripheral circadian clocks which are found in every single cell in the body. Genes that code for proteins that are important for the function of a particular tissue (e.g., liver enzymes in liver cells, neurotransmitters in nerve cells, etc.) show a daily rhythm in gene expression. As a result, all biochemical, physiological and behavioral functions exhibit daily (circadian) rhythms, e.g., body temperature, blood pressure, sleep, cognitive abilities, etc. Notable exceptions are functions that have to be kept within a very narrow range of values, e.g., blood pH and blood concentration of calcium.
So, nervous, endocrine, sensory and circadian systems are all involved in control and regulation of other functions in the body. We will see what happens to all those other functions in the stressed, running zebra next week.
Previously in this series:
BIO101 – Biology and the Scientific Method
BIO101 – Cell Structure
BIO101 – Protein Synthesis: Transcription and Translation
BIO101: Cell-Cell Interactions
BIO101 – From One Cell To Two: Cell Division and DNA Replication
BIO101 – From Two Cells To Many: Cell Differentiation and Embryonic Development
BIO101 – From Genes To Traits: How Genotype Affects Phenotype
BIO101 – From Genes To Species: A Primer on Evolution
BIO101 – What Creatures Do: Animal Behavior
BIO101 – Organisms In Time and Space: Ecology
BIO101 – Origin of Biological Diversity
BIO101 – Evolution of Biological Diversity
BIO101 – Current Biological Diversity
BIO101 – Introduction to Anatomy and Physiology














{kind=link}
{kind=link}